

 22
22 0
0
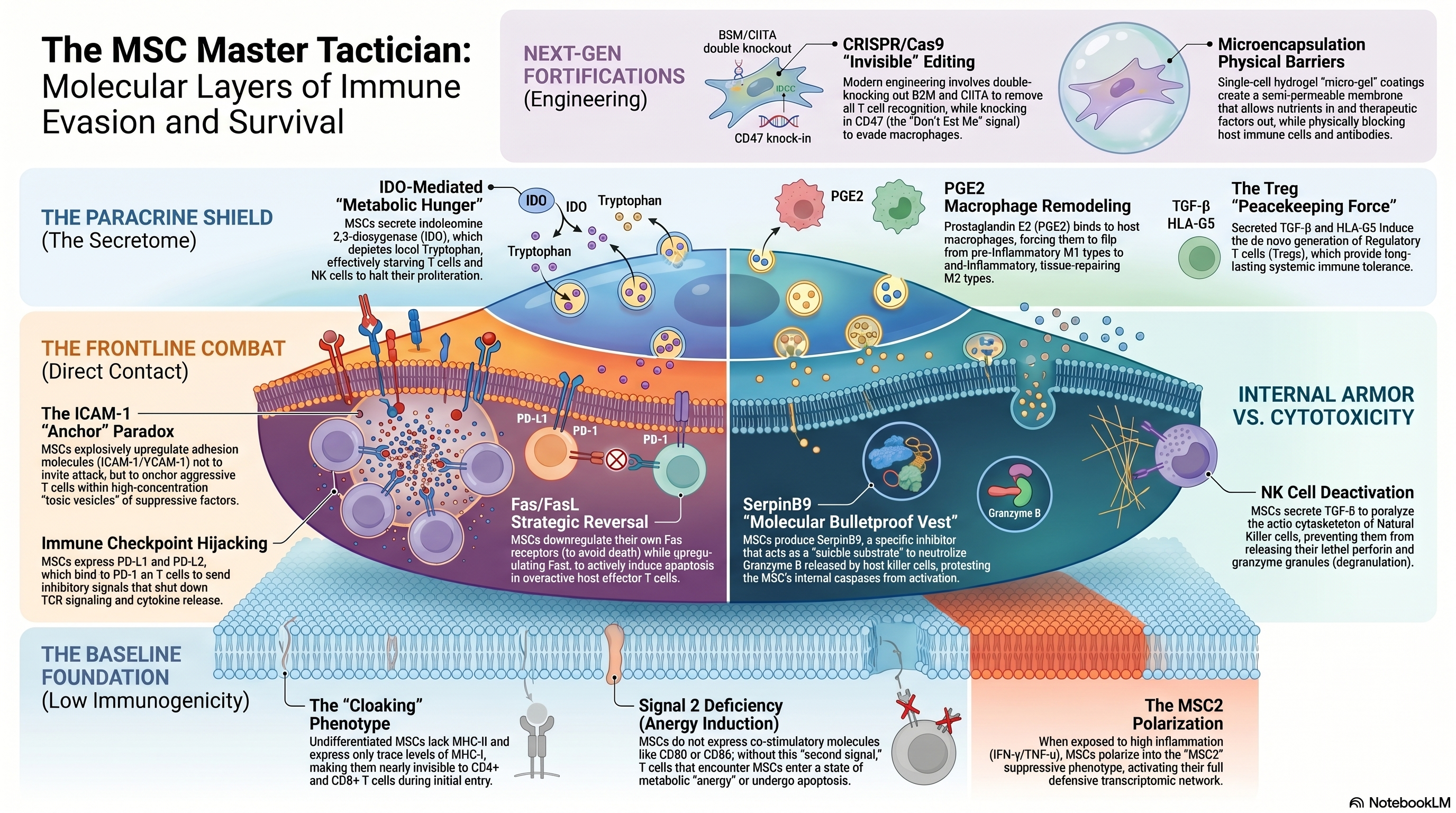
在再生医学早期的叙事里,间充质干细胞(MSCs)一直被披上“免疫特权”的神圣外衣。人们曾乐观地认为,这些细胞如同自带“隐身斗篷”,能跨越遗传屏障在异体内永久定居。
然而,随着高分辨率活体示踪等前沿技术的发展,一场认知范式的转移正在发生:间充质干细胞并非绝对“隐身”,而是一群精通分子博弈的“特工”与“魔术师”。 它们不追求在体内的长生不老,而是通过一系列惊心动魄的“免疫逃逸”战术,在宿主免疫警察的围剿下完成治疗使命。以下是重塑细胞治疗认知的五大真相。
一、 并非“永久定居”,而是“即打即跑”的安全性红利
大众对异体干细胞最大的误区在于“植入即存活”。事实上,异体 MSCs 进入人体后,通常会在 7 到 28 天内被宿主识别并清除。但这并非失败,而是其精妙的**“即打即跑(Hit and Run)”机制**。
MSCs 在消失前,已通过复杂的旁分泌网络深度重塑了宿主的免疫平衡。这种“短暂存续”反而是医学上的安全性红利:正是因为宿主免疫系统最终能够“清理现场”,MSCs 在完成修复任务后会按时离场,从而从底层逻辑上杜绝了外源细胞因不受控增殖而引发肿瘤(致瘤性)的伦理与技术隐忧。
二、 SerpinB9:穿上“分子防弹衣”硬抗剂量博弈
当宿主细胞毒性 T 细胞(CTL)锁定了 MSCs,会释放致命的颗粒酶 B(Granzyme B)。这是一种能切断细胞生命线的“分子手术刀”。然而,MSCs 拥有一种极其罕见的**“分子防弹衣”:SerpinB9 蛋白**。
自杀底物战术: SerpinB9 会伪装成攻击目标,在颗粒酶 B 试图剪切它时,瞬间发生空间构象扭曲,将酶的活性中心彻底锁死。
剂量阈值的博弈: 这种保护并非无限期。SerpinB9 的防御遵循化学计量学原则——它是一种消耗性防御。只有当颗粒酶 B 的浓度低于细胞内储备的 SerpinB9 时,MSCs 才能生存。一旦 CTL 兵临城下释放的毒素超过阈值,防御就会瓦解。正是这种“防御上限”,决定了 MSCs 只能争取到执行任务的宝贵时间窗,而无法永久定居。
三、 空间博弈:将“攻击信号”反转为“抑制锚点”
在传统免疫学中,粘附分子(ICAM-1 和 VCAM-1)是引导免疫细胞进行精准打击的“向导”。但在 MSCs 的世界里,这演变成了一个反直觉的陷阱。
MSCs 会利用这些分子将激活的 T 细胞牢牢“锚定”在自己表面。这看似引火烧身,实则是为了构建一个微米级的“毒性囊泡”微环境。在这个极近距离内,MSCs 释放的免疫抑制因子(如 PGE2)不会被体液稀释,能以超高浓度压制 T 细胞,迫使其陷入免疫无能。

四、 饥饿疗法:IDO 与色氨酸的代谢“焦土政策”
MSCs 还会对免疫细胞实施残酷的“代谢封锁”。通过分泌 IDO(吲哚胺 2,3-双加氧酶),它们在微环境中推行“焦土政策”。
IDO 会快速耗尽免疫细胞生存必需的色氨酸,并产生具有毒性的代谢产物(犬尿氨酸)。对于急需蛋白质合成来扩增“兵力”的效应 T 细胞而言,这无异于粮草断绝。在这种压力下,T 细胞的细胞周期被强制停滞在 G_0/G_1 期。
专家洞察: 值得注意的是,IDO 是人类 MSCs 的核心大杀器;而在小鼠等啮齿类动物中,相应的防御任务则主要由诱导型一氧化氮合酶(iNOS)产生的一氧化氮(NO)来承担。这种物种差异是解读临床前数据的关键。
五、 进化的“通用细胞”:CRISPR 与微囊化的终极伪装
为了让 MSCs 成为“现成可用(Off-the-shelf)”的超级产品,科学家正在对其进行基因组级别的“魔改”:
基因“隐身术”: 利用 CRISPR/Cas9 敲除 B2M(MHC-I 组装的核心)和 CIITA(MHC-II 的转录开关),切断 T 细胞的扫描雷达。
平衡“缺失自我”风险: 单纯隐身会导致 NK 细胞因检测到“缺失自我(Missing Self)”信号而发起疯狂攻击。因此,科学家必须补偿性敲入 CD47(“别吃我”信号)和 HLA-G,通过主动发送抑制信号来平衡由于“隐身”带来的次生风险。
物理“堡垒”: 单细胞微囊化技术。在细胞外包裹一层数微米厚的半透膜“外壳”。这层膜孔径精妙:氧气进得去,因子出得来,但庞大的 CTL 和抗体只能被挡在堡垒之外,实现了物理层面的绝对隔离。
结语:功能性接受的智慧
间充质干细胞的真正智慧在于:它不追求与免疫系统死磕,而是通过精准的代谢干预、空间锚定和分子伪装,在动荡中争取执行使命的时间。
当我们能完美模拟这种“特工”艺术,未来的细胞治疗将像购买抗生素一样便捷,而“排斥反应”也将从医学词典中逐渐淡出。这种从“对抗”到“重塑”的转变,正是再生医学通往未来的终极阶梯。# 细胞界的“特工”与“魔术师”:间充质干细胞免疫逃逸的五大惊人真相
